Abstract
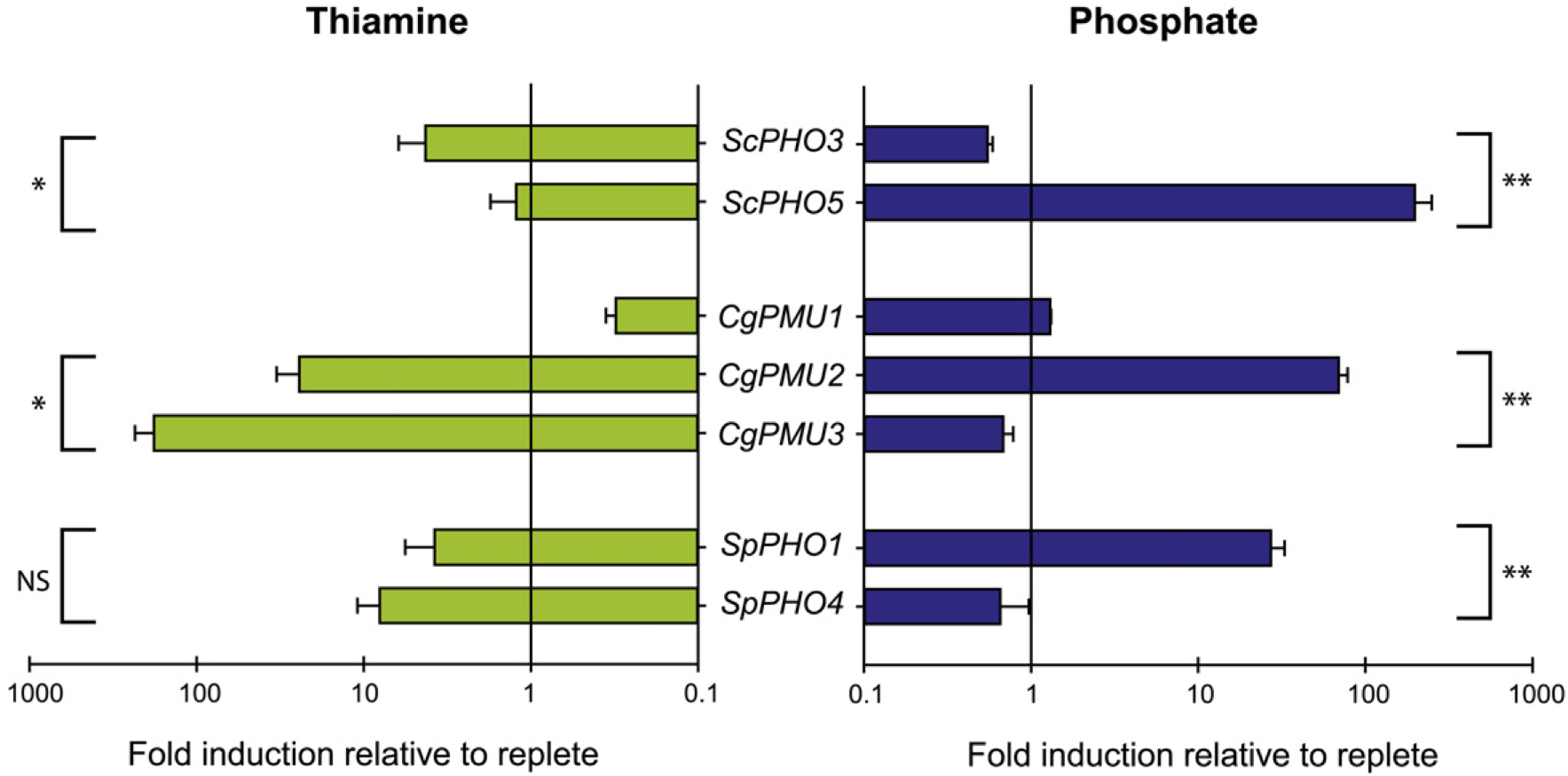
Convergent evolution is often due to selective pressures generating a similar phenotype. We observe relatively recent duplications in a spectrum of Saccharomycetaceae yeast species resulting in multiple phosphatases that are regulated by different nutrient conditions - thiamine and phosphate starvation. This specialization is both transcriptional and at the level of phosphatase substrate specificity. In Candida glabrata, loss of the ancestral phosphatase family was compensated by the co-option of a different histidine phosphatase family with three paralogs. Using RNA-seq and functional assays, we identify one of these paralogs, CgPMU3, as a thiamine phosphatase. We further determine that the 81% identical paralog CgPMU2 does not encode thiamine phosphatase activity; however, both are capable of cleaving the phosphatase substrate, 1-napthyl-phosphate. We functionally demonstrate that members of this family evolved novel enzymatic functions for phosphate and thiamine starvation, and are regulated transcriptionally by either nutrient condition, and observe similar trends in other yeast species. This independent, parallel evolution involving two different families of histidine phosphatases suggests that there were likely similar selective pressures on multiple yeast species to recycle thiamine and phosphate. In this work, we focused on duplication and specialization, but there is also repeated loss of phosphatases, indicating that the expansion and contraction of the phosphatase family is dynamic in many Ascomycetes. The dynamic evolution of the phosphatase gene families is perhaps just one example of how gene duplication, co-option, and transcriptional and functional specialization together allow species to adapt to their environment with existing genetic resources.